
|
Развитие и цветение побегов
|
Про развитие и цветение побегов левзеи сафлоровидной,
связанной с процессом накопления и содержания экдистерона
Продолжение статьи, посвященной технологии получения экдистерон содержащего лекарственного сырья левзеи. Проблемы использования корневищ и корней левзеи для приготовления препаратов экдистерона описаны в статье "Левзея сафлоровидная и препараты на ее основе".
Благодарности. Работа выполнена при частичной финансовой поддержке Администрации Архангельской области и Российского Фонда Фундаментальных Исследований (гранты 03-04-96147, 08-04-98840).
Литературная ссылка. Тимофеев Н.П. Накопление и изменчивость содержания экдистероидов (экдистерона) в лекарственном сырье левзеи сафлоровидной // Сельскохозяйственная биология. Серия “Биология растений”, 2009, № 1. – С. 106-117.
Результаты и их обсуждение: Генеративные побеги левзеи
| 5. Генеративные (репродуктивные) побеги |
Продолжение статьи
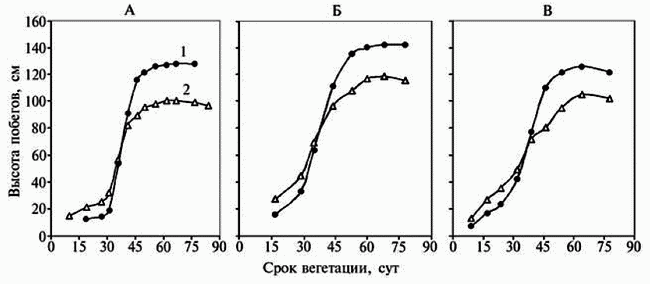
Рис. 2. Динамика роста побегов у растений левзеи Rhaponticum carthamoides в генеративном периоде
(подзона средней тайги европейского Севера): А, Б и В — соответственно 6-й, 7-й и 8-й год жизни, 1 — генеративные, 2 — вегетативные побеги. | |
Продолжение следует на странице: Содержание экдистероидов во фракциях и элементах фитомассы левзеи |
Ссылки на другие публикации из электронных библиотек - в свободном доступе
Про особенности и различия препаратов из Левзеи, Технология Производства и их Активность описаны на следующих страницах нашего сайта "Экдистен, Левзея и Маралий корень: Заказы, Технология, Дозировка и Активность - Ecdystenum, Rhaponticum-Leuzea and Maral root: Orders, Technology, Dosage, Activity":
Экстракт корня Левзеи (экдистерон) или Маральего корня - аналог корня женьшеня, родиолы розовой или золотого корня, китайского лимонника, корня элеуторококка - применяется как биологические добавки к пище, биоактивные добавки, анаболики в бодибилдинге. Экстракт левзеи (или настойка левзеи сафлоровидной) и серпуха используются для снижения артериального давления, также как лекарство от высокого давления, лекарство при гипертонии, входит в препараты от повышенного давления и препараты от высокого давления (полное применение левзеи сафлоровидной). Заказать или купить левзею или семена левзеи сафлоровидной (продажа лекарственных трав) можно на нашем сайте.
Часть 2.
БИОЛОГИЧЕСКАЯ АКТИВНОСТЬ ЭКДИСТЕРОИДОВ, ИХ ИСТОЧНИКОВ И ПРОДУКТОВ
(Включая Экдистен, Экдистерон, Маралий корень, Левзея, Leuzea, Rhaponticum, Maral root)
Часть 3. ЭКДИСТЕН, ЛЕВЗЕЯ-ПОРОШОК И МАРАЛИЙ КОРЕНЬ:
Технология Производства, Химсостав и Дозировка
Купить экдистерона субстанции в Интернет-магазине:
"Левзея +65 экдистеронов и серпуха"